Le mystère de la complexité de la vie
 L'histoire de l'origine de la vie est passionnante, non qu'il n'y ait
plus aucun mystère comme certains le prétendent avec
quelque précipitation, mais au contraire parce qu'elle les
multiplie et dessine un tracé approximatif qui va du chimique au
biologique, des ARN autocatalytiques à la constitution de la
cellule et toute l'évolution qui a suivi jusqu'à nous.
C'est une histoire dont on connaît la fin et les principales
étapes mais où subsistent de multiples
trous plus ou moins obscurs, dont certains d'une importance
considérable. Pourtant, depuis une dizaine d'années on peut
s'enorgueillir d'avoir percé ce qui semblait si obscur
jusqu'ici, et compris les grandes lignes
de l'apparition de la vie sur Terre il y a 3,8 milliards
d'années. Pour la plupart des spécialistes,
désormais l'essentiel est acquis, en fin de compte tout se
réduit à la chimie ! et certes il n'y a plus aucune
nécessité d'un "dessein intelligent" encore moins du
doigt de Dieu dans la création du vivant et de son
évolution. C'est un fait acquis, il faudra que la foi aille se
faire voir ailleurs.
L'histoire de l'origine de la vie est passionnante, non qu'il n'y ait
plus aucun mystère comme certains le prétendent avec
quelque précipitation, mais au contraire parce qu'elle les
multiplie et dessine un tracé approximatif qui va du chimique au
biologique, des ARN autocatalytiques à la constitution de la
cellule et toute l'évolution qui a suivi jusqu'à nous.
C'est une histoire dont on connaît la fin et les principales
étapes mais où subsistent de multiples
trous plus ou moins obscurs, dont certains d'une importance
considérable. Pourtant, depuis une dizaine d'années on peut
s'enorgueillir d'avoir percé ce qui semblait si obscur
jusqu'ici, et compris les grandes lignes
de l'apparition de la vie sur Terre il y a 3,8 milliards
d'années. Pour la plupart des spécialistes,
désormais l'essentiel est acquis, en fin de compte tout se
réduit à la chimie ! et certes il n'y a plus aucune
nécessité d'un "dessein intelligent" encore moins du
doigt de Dieu dans la création du vivant et de son
évolution. C'est un fait acquis, il faudra que la foi aille se
faire voir ailleurs. On
peut regretter la place que prennent les religions
dans le débat
biologique (et le livre de Christian de Duve), y compris dans
l'opposition militante au créationnisme qui va jusqu'à
refuser toute finalité au
vivant, à l'encontre de l'évidence la plus manifeste,
par peur de donner prise au finalisme religieux ! Il y a donc une vue
relativement claire des différents stades de l'émergence
de la vie, mais cela n'empêche pas que leur
interprétation est encore trop incertaine
et réductionniste. La plupart du temps on ne fait rien d'autre
que de se persuader que ce qui s'est passé devait se passer
ainsi (tout ce qui apparaît est bon) ! Il est
très exagéré de prétendre avoir tout
compris ("Aujourd'hui, il n'est pas exagéré d'affirmer que nous connaissons le secret de la vie"
CdD 24) car ce qui est remarquable, c'est que les points d'achoppement,
les trous
de l'histoire, coïncident à chaque fois avec des sauts
inouïs de
complexification. Effectivement, la complexité du fonctionnement des
cellules dépasse l'imagination, chaque processus multipliant les
intermédiaires à foison et l'équilibre entre
réactions contradictoires. Il faut en retenir deux choses :
On
peut regretter la place que prennent les religions
dans le débat
biologique (et le livre de Christian de Duve), y compris dans
l'opposition militante au créationnisme qui va jusqu'à
refuser toute finalité au
vivant, à l'encontre de l'évidence la plus manifeste,
par peur de donner prise au finalisme religieux ! Il y a donc une vue
relativement claire des différents stades de l'émergence
de la vie, mais cela n'empêche pas que leur
interprétation est encore trop incertaine
et réductionniste. La plupart du temps on ne fait rien d'autre
que de se persuader que ce qui s'est passé devait se passer
ainsi (tout ce qui apparaît est bon) ! Il est
très exagéré de prétendre avoir tout
compris ("Aujourd'hui, il n'est pas exagéré d'affirmer que nous connaissons le secret de la vie"
CdD 24) car ce qui est remarquable, c'est que les points d'achoppement,
les trous
de l'histoire, coïncident à chaque fois avec des sauts
inouïs de
complexification. Effectivement, la complexité du fonctionnement des
cellules dépasse l'imagination, chaque processus multipliant les
intermédiaires à foison et l'équilibre entre
réactions contradictoires. Il faut en retenir deux choses : 1. Comme le dit Christian de Duve dans La Recherche, "La direction vers une complexité croissante me paraît un phénomène de stricte observation". John Maynard Smith
n'a sans doute pas tort de répondre à cette constatation qu'"aucune règle de l'évolution ne dit que les choses doivent devenir plus complexes",
car il y a des régressions ou des bactéries n'ayant pas
du tout évolué, ce qui est aussi indéniable.
Certains pensent même que la
cellule originelle était plus complexe que les bactéries
qui en sont issues. Ce n'est pas une raison pour penser que cette
complexité aurait pu apparaître d'un seul coup sans passer par des
stades plus primitifs. Il faut admettre que la complexification est
malgré tout une loi reliée à
celle de
l'entropie, d'ordre statistique elle aussi et fonction du temps
passé. La complexification ne
s'applique pas aux espèces mais seulement à un niveau
plus global où les probabilités s'ajoutent, où les
événements improbables finissent par se produire. Ce
n'est pas une loi qui concerne le court terme, au contraire de
l'entropie cette fois, mais uniquement le long terme.
Au fond la
complexification inverse l'entropie sur le long terme car si le court
terme va presque toujours au plus probable et ramène tout
à la moyenne, plus le temps passe et plus
l'improbable peut se produire et laisser sa trace pour toujours,
organiser
durablement l'espace (c'est une forme d'auto-organisation!). Peu
importe le temps qu'il faudra mais le meilleur l'emportera un jour et
pour toujours même si dans l'immédiat tous les autres
passent devant. La complexification à long terme est tout
simplement la contrepartie du caractère statistique de
l'entropie contre laquelle la vie se défend par le nombre,
abandonnant au vent des semences par milliers.
1. Comme le dit Christian de Duve dans La Recherche, "La direction vers une complexité croissante me paraît un phénomène de stricte observation". John Maynard Smith
n'a sans doute pas tort de répondre à cette constatation qu'"aucune règle de l'évolution ne dit que les choses doivent devenir plus complexes",
car il y a des régressions ou des bactéries n'ayant pas
du tout évolué, ce qui est aussi indéniable.
Certains pensent même que la
cellule originelle était plus complexe que les bactéries
qui en sont issues. Ce n'est pas une raison pour penser que cette
complexité aurait pu apparaître d'un seul coup sans passer par des
stades plus primitifs. Il faut admettre que la complexification est
malgré tout une loi reliée à
celle de
l'entropie, d'ordre statistique elle aussi et fonction du temps
passé. La complexification ne
s'applique pas aux espèces mais seulement à un niveau
plus global où les probabilités s'ajoutent, où les
événements improbables finissent par se produire. Ce
n'est pas une loi qui concerne le court terme, au contraire de
l'entropie cette fois, mais uniquement le long terme.
Au fond la
complexification inverse l'entropie sur le long terme car si le court
terme va presque toujours au plus probable et ramène tout
à la moyenne, plus le temps passe et plus
l'improbable peut se produire et laisser sa trace pour toujours,
organiser
durablement l'espace (c'est une forme d'auto-organisation!). Peu
importe le temps qu'il faudra mais le meilleur l'emportera un jour et
pour toujours même si dans l'immédiat tous les autres
passent devant. La complexification à long terme est tout
simplement la contrepartie du caractère statistique de
l'entropie contre laquelle la vie se défend par le nombre,
abandonnant au vent des semences par milliers.
2. On se heurte à chaque stade à une complexité telle qu'on ne peut en rendre compte au-delà des mécanismes de base et de la sélection naturelle. Ce "mur de la complexité" est peut-être infranchissable, même si on peut considérer que cela n'a pas vraiment d'importance dès lors qu'on tient les deux bouts de la chaîne. Les sauts évolutifs restent pourtant des boîtes noires et notre maîtrise du vivant très approximative. On fait des modèles simplifiés dont le détail nous échappe. La "sélection naturelle" est la formule magique sensée résoudre la question, comme s'il suffisait de secouer la boîte, de faire "abracadabra" et d'attendre un moment pour que tout s'arrange. Le problème, c'est qu'il faudrait attendre très très longtemps ! En fait la place incontournable de la sélection darwinienne dans l'évolution devrait réfuter les prétentions de contrôler le vivant, encore plus de faire mieux ! On peut le réparer voire le modifier mais dans l'optimisation des fonctions rien ne remplace le travail des années.
Il y a beaucoup de choses qu'on sait tout-de-même mais voyons quels sont ces événements improbables qui ponctuent l'histoire de la vie (les transitions de Maynard Smith et Szathmàry) après la formation de la Terre, il y a 4,6 milliards d'années et l'apparition de la vie il y a 3,8 milliards d'année :
1 - L'apparition inexpliquée de molécules réplicantes (ARN auto-catalytique)
2 - Leur confinement dans un compartiment (argile puis membrane), cellule
3 - Le passage de réplicateurs indépendants aux chromosomes qui les regroupe (monde à ARN)
4 - Le passage de l'ARN comme enzyme et gène (LUCA, Last Universal Common Ancestor) à l'ADN (code génétique) et aux protéines (enzymes).
5 - Le passage des procaryotes (bactéries) aux eucaryotes (cellules à noyau), il y a 2,7 milliards d'années (laissant des traces de stéroïdes)
6 - L'apparition de la reproductions sexuée (900 millions ?)
7 - L'apparition des organismes pluricellulaires : plantes, animaux, champignons (différenciation cellulaire) au moins vers 800 millions d'années, avec les premiers bilatériens vers 600 millions d'années (après terre gelée et réchauffement) puis l'explosion du cambrien inaugurant le paléozoïque (542 millions d'années) après de grandes extinctions et une remontée de l'oxygène
8 - La constitution de colonies animales, d'individus sociaux
9 - L'apparition du langage
2 - Leur confinement dans un compartiment (argile puis membrane), cellule
3 - Le passage de réplicateurs indépendants aux chromosomes qui les regroupe (monde à ARN)
4 - Le passage de l'ARN comme enzyme et gène (LUCA, Last Universal Common Ancestor) à l'ADN (code génétique) et aux protéines (enzymes).
5 - Le passage des procaryotes (bactéries) aux eucaryotes (cellules à noyau), il y a 2,7 milliards d'années (laissant des traces de stéroïdes)
6 - L'apparition de la reproductions sexuée (900 millions ?)
7 - L'apparition des organismes pluricellulaires : plantes, animaux, champignons (différenciation cellulaire) au moins vers 800 millions d'années, avec les premiers bilatériens vers 600 millions d'années (après terre gelée et réchauffement) puis l'explosion du cambrien inaugurant le paléozoïque (542 millions d'années) après de grandes extinctions et une remontée de l'oxygène
8 - La constitution de colonies animales, d'individus sociaux
9 - L'apparition du langage

 A partir du quatrième point, c'est l'information qui mène la barque. Les sauts
de complexité les plus importants se situent à
la formation de la cellule sans noyau (bactérienne),
déjà extraordinairement complexe, puis
à la cellule avec noyau (eucaryote), beaucoup plus grosse et
complexe encore. Le passage au stade pluricellulaire serait moins
extraordinaire, s'étant produit plusieurs fois
(végétaux, animaux, champignons), mais il permettra de
faire
éclater cette complexité en une exubérance de
formes taillée à coups de dents par la course entre
prédateurs et proies dans l'espace des possibles, course sans fin entre
contraintes de l'organisme et de l'environnement en co-évolution constante.
A partir du quatrième point, c'est l'information qui mène la barque. Les sauts
de complexité les plus importants se situent à
la formation de la cellule sans noyau (bactérienne),
déjà extraordinairement complexe, puis
à la cellule avec noyau (eucaryote), beaucoup plus grosse et
complexe encore. Le passage au stade pluricellulaire serait moins
extraordinaire, s'étant produit plusieurs fois
(végétaux, animaux, champignons), mais il permettra de
faire
éclater cette complexité en une exubérance de
formes taillée à coups de dents par la course entre
prédateurs et proies dans l'espace des possibles, course sans fin entre
contraintes de l'organisme et de l'environnement en co-évolution constante.La reproduction et l'information
Qu'est-ce que la vie ? Elle résulte de processus chimiques complexes où interviennent principalement trois types de molécules géantes capables de transmettre de l'information : les protéines et deux familles d'acides nucléiques, l'acide ribonucléique (ARN) et l'acide désoxyribonucléique (ADN). La Recherche p34
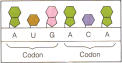 L'évolution
est une diversification et une complexification mais, avant tout
organisme complexe, il faut une solide reproduction de l'information.
Tout découle de la reproduction
qui
ajoute à la
chimie l'information, aussi bien dans la suite d'acides aminés
des protéines que dans la suite des gènes de l'ADN
combinant 4 bases : Adénine, Guanine, Cytosine, Thymine (ou
Uracile pour l'ARN) désignés par leur initiale (AGCT). En
fait on peut dire que la vie, ce n'est pas
autre chose que la reproduction elle-même. C'est ce qui fait que
dès qu'une chimie de la reproduction se met en route
(autocatalyse de l'ARN), on quitte le domaine de la chimie pour celui
de la "sélection darwinienne", c'est-à-dire de la
sélection par le résultat, par rétroaction
extérieure, par "causalité descendante" et non plus par
causalité chimique : seul ce qui se reproduit le mieux continue
à se reproduire, même s'il est très improbable au
départ, à cause de son amplification ultérieure
par la reproduction justement. L'évolution procède en
effet par
pression adaptative, mutation,
sélection, et amplification (reproduction). Il faut d'ailleurs
distinguer les mutations majeures, qui apportent de nouvelles fonctions
et changent d'environnement, des mutations adaptatives qui vont suivre.
L'évolution
est une diversification et une complexification mais, avant tout
organisme complexe, il faut une solide reproduction de l'information.
Tout découle de la reproduction
qui
ajoute à la
chimie l'information, aussi bien dans la suite d'acides aminés
des protéines que dans la suite des gènes de l'ADN
combinant 4 bases : Adénine, Guanine, Cytosine, Thymine (ou
Uracile pour l'ARN) désignés par leur initiale (AGCT). En
fait on peut dire que la vie, ce n'est pas
autre chose que la reproduction elle-même. C'est ce qui fait que
dès qu'une chimie de la reproduction se met en route
(autocatalyse de l'ARN), on quitte le domaine de la chimie pour celui
de la "sélection darwinienne", c'est-à-dire de la
sélection par le résultat, par rétroaction
extérieure, par "causalité descendante" et non plus par
causalité chimique : seul ce qui se reproduit le mieux continue
à se reproduire, même s'il est très improbable au
départ, à cause de son amplification ultérieure
par la reproduction justement. L'évolution procède en
effet par
pression adaptative, mutation,
sélection, et amplification (reproduction). Il faut d'ailleurs
distinguer les mutations majeures, qui apportent de nouvelles fonctions
et changent d'environnement, des mutations adaptatives qui vont suivre.La vie (ou l'évolution) naît de la reproduction, quand la chimie est sous contrainte et n'est plus déterminante mais étroitement contrôlée par l'information qui émerge du processus par rétroaction après un temps plus ou moins long de reproduction et de sélection. On peut dire que l'information comme mémoire sélective résulte d'une optimisation de la réaction, et donc de la reproduction finale. Cette inversion de causalité est le coeur du darwinisme, toujours aussi difficile à comprendre pour le réductionnisme car le déterminisme devient extérieur. En effet, la causalité vient désormais de l'environnement qui n'a de sens que pour un organisme vivant, c'est-à-dire fait pour durer et se reproduire dans un espace limité et un temps qui reste imprévisible. Dès lors, il vaut mieux dans le court terme ne pas occuper tout l'espace (être trop bien adapté) afin de garder ses capacités d'adaptation et de reproduction à plus long terme. Car le temps long est une dimension fondamentale de la vie et de son inversion de l'entropie au travers de toutes sortes de cataclysmes et d'extinctions de masse. Les phénomènes biologiques relèvent d'abord de la loi des grand nombres et de temps géologiques.
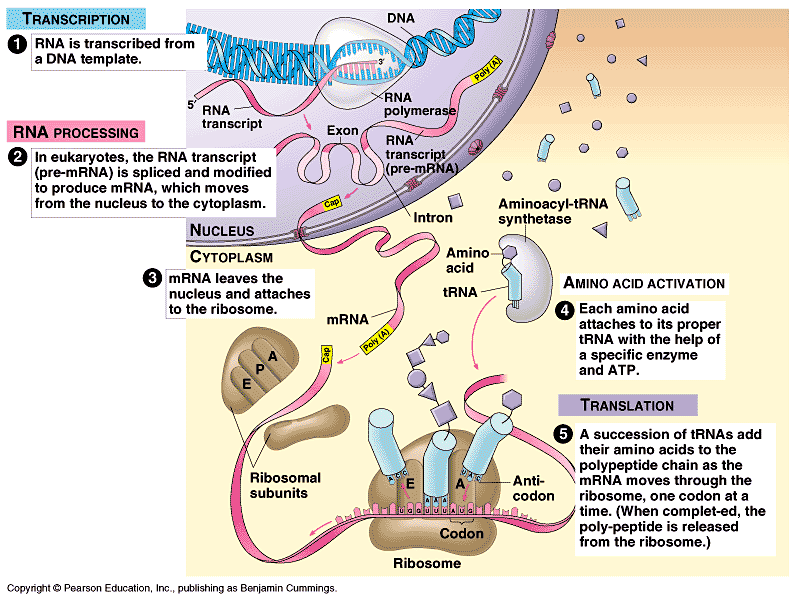 Comment
peut-on décrire la vie d'une cellule de
l'intérieur ? C'est
essentiellement
une production incessante, par l'ARN à partir de l'ADN, de
protéines, molécules
géantes se
repliant en formes
complexes et qui ont des fonctions catalytiques (enzymes), participant
au métabolisme de la cellule (constitué du catabolisme
destructeur et de l'anabolisme constructeur), notamment les divers
cycles du cytoplasme qui
maintiennent l'homéostasie (les cycles permettent de
rétablir alternativement les excès et les manques de
l'équilibre interne). A la base de cette énergie vitale,
il y a une horloge biologique dont le principe est très simple :
Un gène produit une ou plusieurs protéines (PER/CRY) qui l'inhibe jusqu'à
ce que la protéine se décompose réactivant le
gène qui la produit à nouveau, etc. Le temps de
décomposition de cette protéine est donc l'unité
de base d'une horloge interne sur laquelle de nombreux processus vont
se greffer, dans une programmation complexe des cycles biologiques. Ce n'est pas si
différent d'un ordinateur dont la cadence d'horloge détermine le
temps de traitement de l'information.
L'énergie vitale de
la cellule est catalytique, c'est une chaîne de réactions
qui s'engendrent constamment. A ce sujet, il est intéressant de préciser que certains vers microscopiques (tardigrades)
peuvent s'arrêter de vivre en séchant (cryptobiose), se
conserver ainsi indéfiniment, puis reprendre vie avec un peu d'eau
comme des graines !
Comment
peut-on décrire la vie d'une cellule de
l'intérieur ? C'est
essentiellement
une production incessante, par l'ARN à partir de l'ADN, de
protéines, molécules
géantes se
repliant en formes
complexes et qui ont des fonctions catalytiques (enzymes), participant
au métabolisme de la cellule (constitué du catabolisme
destructeur et de l'anabolisme constructeur), notamment les divers
cycles du cytoplasme qui
maintiennent l'homéostasie (les cycles permettent de
rétablir alternativement les excès et les manques de
l'équilibre interne). A la base de cette énergie vitale,
il y a une horloge biologique dont le principe est très simple :
Un gène produit une ou plusieurs protéines (PER/CRY) qui l'inhibe jusqu'à
ce que la protéine se décompose réactivant le
gène qui la produit à nouveau, etc. Le temps de
décomposition de cette protéine est donc l'unité
de base d'une horloge interne sur laquelle de nombreux processus vont
se greffer, dans une programmation complexe des cycles biologiques. Ce n'est pas si
différent d'un ordinateur dont la cadence d'horloge détermine le
temps de traitement de l'information.
L'énergie vitale de
la cellule est catalytique, c'est une chaîne de réactions
qui s'engendrent constamment. A ce sujet, il est intéressant de préciser que certains vers microscopiques (tardigrades)
peuvent s'arrêter de vivre en séchant (cryptobiose), se
conserver ainsi indéfiniment, puis reprendre vie avec un peu d'eau
comme des graines !{kind=link}
 La
fonction d'horloge de base, ou d'énergie vitale, pourrait
d'ailleurs s'appliquer de façon plus pertinente encore au
déplacement incessant de l'ARN-polymérase le long de
l'ADN, effectuant la transcription de l'ARN-messager qui sera traduit
ensuite en protéine dans le ribosome, s'il ne pouvait y avoir plusieurs
ARN-polymérase décodant en même temps des parties
différentes de l'ADN. L'intéressant ici,
c'est la sélection des parties à transcrire, des
commandes à effectuer, car c'est une toute petite partie de
l'ADN qui sera lue à chaque fois, même si cela
représente déjà des milliers de base. Ainsi chaque
cellule de notre
corps possède tout notre matériel génétique
mais seule une partie des gènes est activée dans chacune,
selon sa fonction, par des "facteurs d'expression", protéines
produites par des gènes régulateurs (ou
"supergènes"). Le mécanisme d'expression des
gènes,
découvert par Jacob et Monod, (Nobels 1965) ressemble au
mécanisme d'horloge décrit plus haut dont le principe est
significativement
celui d'une levée d'inhibition.
En effet, un
"opéron" (gène opératoire) commence normalement
par un code
de départ, codon-start dit "promoteur" (le plus souvent, pour
les
eucaryotes, TATAAT, appelé TATA box, alternance de
Thymine et Adénine, mais qui peut être parfois ATG entre
autres). C'est l'équivalent d'un
X-on en
informatique. L'opéron se termine aussi par une
séquence terminale de 3
codons-stop (UAG, UAA et UGA), équivalent d'un X-off. Or, le
code de
départ se trouve la plupart du temps inhibé par un
"répresseur"
spécifique qui le rend illisible et s'applique sur un ensemble
fonctionnel de gènes. Il
faut que surgisse un "inducteur" venant se fixer sur le
répresseur pour que celui-ci lâche prise et libère
la lecture du codon-start, ce qui enclenche
la
production de la protéine codée par le gène
concerné.
L'inducteur, le plus souvent signal d'un déséquilibre
à réduire, représente l'information circulante,
une perception
extérieure codifiée, une commande externe.
L'opéron qui y répond
est
l'information structure, l'organisation matérielle, la
production de protéines en cascade, la
procédure interne mémorisée.
La
fonction d'horloge de base, ou d'énergie vitale, pourrait
d'ailleurs s'appliquer de façon plus pertinente encore au
déplacement incessant de l'ARN-polymérase le long de
l'ADN, effectuant la transcription de l'ARN-messager qui sera traduit
ensuite en protéine dans le ribosome, s'il ne pouvait y avoir plusieurs
ARN-polymérase décodant en même temps des parties
différentes de l'ADN. L'intéressant ici,
c'est la sélection des parties à transcrire, des
commandes à effectuer, car c'est une toute petite partie de
l'ADN qui sera lue à chaque fois, même si cela
représente déjà des milliers de base. Ainsi chaque
cellule de notre
corps possède tout notre matériel génétique
mais seule une partie des gènes est activée dans chacune,
selon sa fonction, par des "facteurs d'expression", protéines
produites par des gènes régulateurs (ou
"supergènes"). Le mécanisme d'expression des
gènes,
découvert par Jacob et Monod, (Nobels 1965) ressemble au
mécanisme d'horloge décrit plus haut dont le principe est
significativement
celui d'une levée d'inhibition.
En effet, un
"opéron" (gène opératoire) commence normalement
par un code
de départ, codon-start dit "promoteur" (le plus souvent, pour
les
eucaryotes, TATAAT, appelé TATA box, alternance de
Thymine et Adénine, mais qui peut être parfois ATG entre
autres). C'est l'équivalent d'un
X-on en
informatique. L'opéron se termine aussi par une
séquence terminale de 3
codons-stop (UAG, UAA et UGA), équivalent d'un X-off. Or, le
code de
départ se trouve la plupart du temps inhibé par un
"répresseur"
spécifique qui le rend illisible et s'applique sur un ensemble
fonctionnel de gènes. Il
faut que surgisse un "inducteur" venant se fixer sur le
répresseur pour que celui-ci lâche prise et libère
la lecture du codon-start, ce qui enclenche
la
production de la protéine codée par le gène
concerné.
L'inducteur, le plus souvent signal d'un déséquilibre
à réduire, représente l'information circulante,
une perception
extérieure codifiée, une commande externe.
L'opéron qui y répond
est
l'information structure, l'organisation matérielle, la
production de protéines en cascade, la
procédure interne mémorisée. L'énergie chimique
est
stockée sous forme d'ATP
(adénosine
triphosphate) qui fournit de l'énergie en se réduisant en
ADP
(adénosine diphosphate) grâce à des
"transducteurs". Il faut souligner que l'ATP lorsqu'il perd 2
phosphates sur 3 n'est
rien d'autre que l'Adénine et qu'on trouve aussi, bien que
beaucoup moins, CTP, GTP et UTP basés sur les autres acides
nucléiques. Dans la cellule, énergie et information sont
donc reliés. L'énergie (ATP) précède
l'information (Adénine) mais si l'information est
inactive par elle-même, l'énergie est bien guidée par l'information
puisqu'elle est au service de la
production de protéines. On peut dire qu'elle alimente les
catalyses qui sont le
véritable
moteur, les
protéines produites servant ensuite de "pompes"
à travers
la
membrane de molécules spécifiques aussi
bien que d'évacuation, voire de toxine contre un agresseur,
assurant
l'homéostasie interne, la défense et la reproduction de
la cellule (méiose et mitose). Il doit être clair que ce
qui organise la cellule, ce n'est pas la
complexité des cascades de réactions chimiques,
sélectionnées à la longue, mais bien la fonction,
l'avantage reproductif (et ce n'est évidemment pas le
gène qui se reproduit, hypothèse imbécile, mais la
fonction qu'il permet avec l'introduction de cette protéine, sa
justesse, son optimisation, la vérité qu'il dit sur le
monde, une sorte de mémoire perceptive, de rémanence du
vivant, de caractère acquis au cours de l'évolution,
sélectionné par sa
fonction, même si c'est bien par le gène que tout commence
et qu'il y a des mutations neutres ou d'abord inutiles). "Un
stade doit nécessairement arriver où la sélection
des molécules ne se fonde plus sur ce qu'elles sont, mais sur ce
qu'elles font" CdD p107.
L'énergie chimique
est
stockée sous forme d'ATP
(adénosine
triphosphate) qui fournit de l'énergie en se réduisant en
ADP
(adénosine diphosphate) grâce à des
"transducteurs". Il faut souligner que l'ATP lorsqu'il perd 2
phosphates sur 3 n'est
rien d'autre que l'Adénine et qu'on trouve aussi, bien que
beaucoup moins, CTP, GTP et UTP basés sur les autres acides
nucléiques. Dans la cellule, énergie et information sont
donc reliés. L'énergie (ATP) précède
l'information (Adénine) mais si l'information est
inactive par elle-même, l'énergie est bien guidée par l'information
puisqu'elle est au service de la
production de protéines. On peut dire qu'elle alimente les
catalyses qui sont le
véritable
moteur, les
protéines produites servant ensuite de "pompes"
à travers
la
membrane de molécules spécifiques aussi
bien que d'évacuation, voire de toxine contre un agresseur,
assurant
l'homéostasie interne, la défense et la reproduction de
la cellule (méiose et mitose). Il doit être clair que ce
qui organise la cellule, ce n'est pas la
complexité des cascades de réactions chimiques,
sélectionnées à la longue, mais bien la fonction,
l'avantage reproductif (et ce n'est évidemment pas le
gène qui se reproduit, hypothèse imbécile, mais la
fonction qu'il permet avec l'introduction de cette protéine, sa
justesse, son optimisation, la vérité qu'il dit sur le
monde, une sorte de mémoire perceptive, de rémanence du
vivant, de caractère acquis au cours de l'évolution,
sélectionné par sa
fonction, même si c'est bien par le gène que tout commence
et qu'il y a des mutations neutres ou d'abord inutiles). "Un
stade doit nécessairement arriver où la sélection
des molécules ne se fonde plus sur ce qu'elles sont, mais sur ce
qu'elles font" CdD p107.L'évolution renforce le rôle de l'information au fur et à mesure de ces reproductions et sélections qui ne mènent pas à l'uniformité mais à la complexification, la diversification, la différenciation pour faire face à l'imprévisibilité du monde. Le caractère de totalité du vivant (de clôture opérationnelle) va se renforcer dans le même mouvement, de la membrane cellulaire au noyau central, puis totalité sexuelle, totalité d'un organisme, totalité sociale, langage...
 Il est remarquable que l'avantage reproductif de la sexualité
n'ait absolument rien d'évident pour les spécialistes puisque le coût reproductif des mâles (qui
n'enfantent pas) est la division par 2 de la fécondité
à chaque génération ! On s'imagine que c'est le
mélange des gènes qui expliquerait son succès
adaptatif mais c'est contredit par le fait que les bactéries
échangent souvent leurs gènes ("c'est le voisinage écologique des partenaires qui favorise les échanges"
p44). Le véritable
avantage me semble plutôt à chercher du côté
de la communication qui doit s'élaborer entre sexes, sur la
sélection qui s'opère par le choix
du partenaire, du partage d'un code commun, d'une capacité de
coopération et de réactivité commune à
l'environnement, un début de solidarité de groupe et de
normalisation. De plus la sexualité introduit un certain
degré de divergence génétique (dimorphisme sexuel)
permettant une division des fonctions et donc leur optimisation,
véritable complémentarité.
Il est remarquable que l'avantage reproductif de la sexualité
n'ait absolument rien d'évident pour les spécialistes puisque le coût reproductif des mâles (qui
n'enfantent pas) est la division par 2 de la fécondité
à chaque génération ! On s'imagine que c'est le
mélange des gènes qui expliquerait son succès
adaptatif mais c'est contredit par le fait que les bactéries
échangent souvent leurs gènes ("c'est le voisinage écologique des partenaires qui favorise les échanges"
p44). Le véritable
avantage me semble plutôt à chercher du côté
de la communication qui doit s'élaborer entre sexes, sur la
sélection qui s'opère par le choix
du partenaire, du partage d'un code commun, d'une capacité de
coopération et de réactivité commune à
l'environnement, un début de solidarité de groupe et de
normalisation. De plus la sexualité introduit un certain
degré de divergence génétique (dimorphisme sexuel)
permettant une division des fonctions et donc leur optimisation,
véritable complémentarité.La complémentarité
Le principe de base de l'information biochimique et de la reproduction, c'est la complémentarité, ce qu'on appelle l'appariement des bases qui permet la transcription de l'ADN à l'ARN puis la traduction de l'ARN en protéines (mais c'est aussi le principe des récepteurs des cellules avec les neuromédiateurs par exemple, sans parler de la sexualité) :
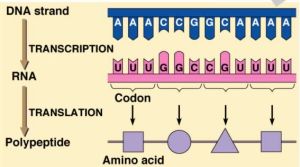
 -
Les acides nucléiques(ARN
et ADN) se caractérisent
par
l'association d'un sucre (ribose pour l'ARN, désoxyribose pour
l'ADN), d'un acide phosphorique et d'une base azotée. Ils se
lient en longues chaînes. La transcription de ces longues
chaînes se fait par un
système de moule, ou de négatif plutôt puisque les
codons seront traduits en anti-codons par l'ARN avant de servir
à sélectionner les acides aminés dans les
protéines ou recopier un brin d'ADN. L'ADN est une sorte d'ARN
désactivé,
très peu réactif et plus rigide, une mémoire
inerte. L'ARN plus réactif et fragile, est devenu
son messager. Tout ceci n'est possible qu'à disposer d'un nombre
réduit de bases (A, G, C, T/U) pouvant se combiner 2 à 2
(A-U ou G-C) et constituant un code correspondant à des acides
aminés, ce n'est
pas rien. Ainsi un ARN porteur uniquement d'Uracile produit de la
phenylalanine (Nirenberg, 1961). La complémentarité est
aussi ce qui explique les deux brins d'ADN permettant une reproduction
presque parfaite en assurant une redondance de l'information qui permet
de contrôler sa validité (mécanisme de correction
d'erreur). Le mécanisme de reproduction est si parfait qu'il
produit un peu moins d'une erreur sur 1 milliard ! Cependant "remarquons
que cela fait encore une demi-douzaine d'erreurs chaque fois qu'une
cellule humaine recopie son génome avant de se diviser" CdD p54
-
Les acides nucléiques(ARN
et ADN) se caractérisent
par
l'association d'un sucre (ribose pour l'ARN, désoxyribose pour
l'ADN), d'un acide phosphorique et d'une base azotée. Ils se
lient en longues chaînes. La transcription de ces longues
chaînes se fait par un
système de moule, ou de négatif plutôt puisque les
codons seront traduits en anti-codons par l'ARN avant de servir
à sélectionner les acides aminés dans les
protéines ou recopier un brin d'ADN. L'ADN est une sorte d'ARN
désactivé,
très peu réactif et plus rigide, une mémoire
inerte. L'ARN plus réactif et fragile, est devenu
son messager. Tout ceci n'est possible qu'à disposer d'un nombre
réduit de bases (A, G, C, T/U) pouvant se combiner 2 à 2
(A-U ou G-C) et constituant un code correspondant à des acides
aminés, ce n'est
pas rien. Ainsi un ARN porteur uniquement d'Uracile produit de la
phenylalanine (Nirenberg, 1961). La complémentarité est
aussi ce qui explique les deux brins d'ADN permettant une reproduction
presque parfaite en assurant une redondance de l'information qui permet
de contrôler sa validité (mécanisme de correction
d'erreur). Le mécanisme de reproduction est si parfait qu'il
produit un peu moins d'une erreur sur 1 milliard ! Cependant "remarquons
que cela fait encore une demi-douzaine d'erreurs chaque fois qu'une
cellule humaine recopie son génome avant de se diviser" CdD p54 - Les acides aminés,
qui sont les briques
élémentaires des protéines,
présents dans la "soupe primitive" (Stanley Miller, 1953), se
caractérisent par
un groupement
acide et un groupement amine, ainsi qu'un autre groupement
différent
pour les 20 acides aminés qui constituent l'alphabet
utilisé par toutes les protéines dans les cellules
vivantes (sous leur forme, on dit leur chiralité, tournée
vers
la gauche à l'exclusion de la droite tout aussi courante
chimiquement,
sans doute pour ne pas mélanger les formes). La liaison du
groupement
acide au groupement amine d'un autre
acide
aminé constitue une "liaison peptidique", principe d'assemblage
de
longues chaînes d'acides aminés (par les ribosomes,
qui sont des
ARN servant d'enzymes et qui ajoutent des séquences
d'acides aminés aux protéines).
- Les acides aminés,
qui sont les briques
élémentaires des protéines,
présents dans la "soupe primitive" (Stanley Miller, 1953), se
caractérisent par
un groupement
acide et un groupement amine, ainsi qu'un autre groupement
différent
pour les 20 acides aminés qui constituent l'alphabet
utilisé par toutes les protéines dans les cellules
vivantes (sous leur forme, on dit leur chiralité, tournée
vers
la gauche à l'exclusion de la droite tout aussi courante
chimiquement,
sans doute pour ne pas mélanger les formes). La liaison du
groupement
acide au groupement amine d'un autre
acide
aminé constitue une "liaison peptidique", principe d'assemblage
de
longues chaînes d'acides aminés (par les ribosomes,
qui sont des
ARN servant d'enzymes et qui ajoutent des séquences
d'acides aminés aux protéines).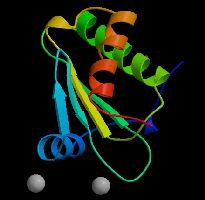 C'est ainsi que se forme
une
chaîne de polypeptides, de centaines ou
milliers d'acides aminés, qu'on appelle une
protéine, véritable machinerie biologique. En effet,
l'interaction entre les liaisons peptidiques replie la
protéine en hélice. L'interaction entre les
différents radicaux lui donne une forme tridimensionnelle. Enfin
l'interaction entre les différentes unités lui donne une
conformation globulaire, spiralée, etc. On peut dire que c'est
cette forme qui est
codée dans une séquence de gènes et qui
détermine ses
propriétés (c'est une structure dynamique en mouvement,
une véritable machine). Chaque protéine "reconnaît"
spécifiquement une ou plusieurs molécules
selon un principe de complémentarité de type
clé-serrure, grâce à des
sites de reconnaissance et de fixation situés à sa
surface. La biochimie, basée sur le carbone, est une chimie des formes, une
chimie in-formée par l'ADN qui est la mémoire de la vie.
C'est ainsi que se forme
une
chaîne de polypeptides, de centaines ou
milliers d'acides aminés, qu'on appelle une
protéine, véritable machinerie biologique. En effet,
l'interaction entre les liaisons peptidiques replie la
protéine en hélice. L'interaction entre les
différents radicaux lui donne une forme tridimensionnelle. Enfin
l'interaction entre les différentes unités lui donne une
conformation globulaire, spiralée, etc. On peut dire que c'est
cette forme qui est
codée dans une séquence de gènes et qui
détermine ses
propriétés (c'est une structure dynamique en mouvement,
une véritable machine). Chaque protéine "reconnaît"
spécifiquement une ou plusieurs molécules
selon un principe de complémentarité de type
clé-serrure, grâce à des
sites de reconnaissance et de fixation situés à sa
surface. La biochimie, basée sur le carbone, est une chimie des formes, une
chimie in-formée par l'ADN qui est la mémoire de la vie. 
Liaison peptidique
La sainte trinité
Le vivant sait compter jusqu'à 3. Il n'y a pas que l'unité primordiale de la cellule et la complémentarité 2 à 2 dans la vie car on retrouve des structures ternaires à différents niveaux:
- L'assemblage des acides aminés se fait par groupes de 3, correspondant aux triplets de bases de l'ADN appelés codons. Si les bases sont des bits qui peuvent avoir des valeurs de 1 à 4, les codons sont des bytes (ou octets) constitués de 3 bases permettant de coder 64 acides aminés différents (même s'il n'y en a que 20 qui marchent et sont traduits par des ARN de transfert).
- Les protéines correspondent aux fonctions et caractéristiques d'une cellule, y compris sa reproduction. Elles font partie d'un système triple ADN/ARN/protéines où chaque partie dépend des autres. L'ADN contient l'information génétique de base (codons), l'ARN les traduits en protéines, les protéines assurent la reproduction et l'homéostasie de l'ensemble. On a donc séparation entre mémoire, lecture (commande) et fonction. L'ARN d'où vient la vie n'est plus que son médiateur qui se sert de moins réactif que lui (ADN) pour produire plus réactif que lui (protéine).
- Il y a aussi 3 sortes de cellules : bactéries, archéobactéries (ou archées) et cellules eucaryotes. Ici, on a sans doute d'abord 2 sortes d'ADN, puis une cellule qui réunit les deux en symbiose puisque le noyau de la cellule eucaryote serait à l'origine une archéobactérie intégrée dans une bactérie (ou le contraire, ou un virus).
- Il y a enfin 3 sortes d'organismes pluricellulaires. Pourquoi 3 ? Sans doute les champignons (levures, moisissures) sont une forme intermédiaire, proche des cellules animales (de la chair plus que de la cellulose), et qui vivent en parasite ou symbiose d'autres organismes leurs fournissant énergie et carbone. La division entre végétal et animal oppose plante "autotrophe", qui trouve son énergie directement dans la matière inanimée (photosynthétiques ou chimiosynthétiques), et l'animal hétérotrophe qui retire son énergie chimique d'autres matières organiques : tous les animaux sont des prédateurs, même les herbivores, et toutes les plantes sont des capteurs d'énergie. Les champignons sont au milieu, "absorbotrophes", plutôt du côté animal mais sans vouloir la mort de ceux qui les nourrissent, en parasites ou en symbiose (lichens), apportant minéraux et protection bactérienne à leurs hôtes. Leur rôle d'équilibre bactérien est sans doute ce qui leur procure un espace vital irremplaçable.
On voit que la vie incarne 4 sortes de
dialectiques
: celle qui de 3 fait 1, celle qui fait du tiers
l'entre-deux, le médiateur, celle qui fait un troisième
de la réunion des deux, enfin celle de la voie moyenne, du
compromis. Le rôle du
troisième n'est pas le même à chaque fois. Le
premier est quelconque, partie d'un tout, le second est liaison active
entre l'un et l'autre, le suivant intègre l'un dans l'autre, le
dernier est composé, mélange, ni l'un, ni l'autre, au milieu. On
occuperait ainsi tout l'espace des
possibles ? Mais voyons comment ça c'est passé, du moins
ce qu'on croit en savoir à ce jour.
La naissance de la vie (reconstitution)
L'ARN est une molécule déjà très complexe, qui n'a jamais été synthétisée en laboratoire sans enzymes. La plupart des chercheurs pensent aujourd'hui que l'apparition de la vie est antérieure à celle de l'ARN. La Recherche, p35

L'origine nous échappe toujours. Nous ne savons pas ce qui a précédé l'ARN, nous savons que la vie a commencé avec des molécules d'ARN autoréplicatives (Walter Gilbert 1986), peut-être elles-mêmes le produit d'une autre molécule autoréplicante (acide aminés "activés", soufrés, ou thioesters en chaînes peptidiques, qu'on trouve dans les cheminées hydrothermales sous-marines), mais ce serait encore un miracle que de l'ARN se forme spontanément, même si ce n'est pas tout-à-fait de l'ordre de l'impossible (à partir d'une réaction dite de formose qui consiste à polymériser les molécules de formaldéhyde en longue chaîne. D'autres hypothèses partent du monoxyde d'azote). En tout cas, il est certain que ce devait être au début des petits ARN (dits ARN interférents) constitués d'une vingtaine d'acides nucléiques seulement (quoique la capacité catalytique soit reliée à la capacité de se plier de l'ARN qui doit donc être assez long pour cela). On peut considérer qu'à partir de là tout est joué, mais comment y parvenir sinon déjà par une reproduction sélective ? (Ce que Antoine Danchin appelle une chimie sélective, aboutissant selon lui à l'auto-réplication de ptéridine-phosphate ou autres peptides). Hélas, il n'est pas possible de remonter au-delà de la mémoire du vivant, fixée dans l'ARN jusqu'à nos jours. En tout cas, ce qu'il faut au début de la vie, c'est l'emballement d'un mécanisme de reproduction chimique plus simple et d'abord sans mémoire, véritable réaction en chaîne qui dure encore. On ne peut dire qu'on sait vraiment ce qui a mis en mouvement ce monde de la vie, pas plus d'ailleurs qu'on ne connaît la cause du Big Bang à l'origine de notre univers. Pas de quoi donner à quelque dieu ce petit rôle de starter (de premier moteur d'Aristote qui enclenche le mouvement) mais il faut reconnaître que la cause première reste hors de notre portée, l'étincelle initiale (il y en a eu d'innombrables sans doute).
 La
reproduction et la sélection de l'ARN
autocatalytique a peut-être déjà commencé
à constituer un proto-métabolisme dans les pores de l'argile. La prochaine étape qui tient encore
du miracle, c'est la constitution d'une membrane,
du moins son
intégration au matériel génétique
porté par l'ARN. Bien sûr les membranes n'ont rien de
mystérieux. Ce sont des molécules phospho-lipidiques
(acides gras) dont un
côté est hydrophile et l'autre hydrophobe, ce sont des
petites bulles huileuses comme il y en a tant, mais on ne sait comment
ça s'est produit.
Il est possible que les membranes lipidiques aient été
précédées de membranes protéinoïdes qui se
produisent quand on chauffe des acides aminés. Selon Christian
de Duve il n'est pas nécessaire que la membrane soit
intégrée au code génétique car toute
membrane vient d'une membrane et peut donc avoir une vie autonome.
Remarquons qu'il
n'est pas impossible que proto-métabolisme et
membrane soient antérieurs à l'ARN, et même que
tout ait commencé par la reproduction de membranes (c'est l'hypothèse d'Antoine Danchin mais semble contredit par
le fait que bactéries et archéobactéries n'ont pas
les mêmes membranes).
La
reproduction et la sélection de l'ARN
autocatalytique a peut-être déjà commencé
à constituer un proto-métabolisme dans les pores de l'argile. La prochaine étape qui tient encore
du miracle, c'est la constitution d'une membrane,
du moins son
intégration au matériel génétique
porté par l'ARN. Bien sûr les membranes n'ont rien de
mystérieux. Ce sont des molécules phospho-lipidiques
(acides gras) dont un
côté est hydrophile et l'autre hydrophobe, ce sont des
petites bulles huileuses comme il y en a tant, mais on ne sait comment
ça s'est produit.
Il est possible que les membranes lipidiques aient été
précédées de membranes protéinoïdes qui se
produisent quand on chauffe des acides aminés. Selon Christian
de Duve il n'est pas nécessaire que la membrane soit
intégrée au code génétique car toute
membrane vient d'une membrane et peut donc avoir une vie autonome.
Remarquons qu'il
n'est pas impossible que proto-métabolisme et
membrane soient antérieurs à l'ARN, et même que
tout ait commencé par la reproduction de membranes (c'est l'hypothèse d'Antoine Danchin mais semble contredit par
le fait que bactéries et archéobactéries n'ont pas
les mêmes membranes).Le rassemblement de tout le matériel génétique en chromosomes rassemblant les ARN codants et représentant des centaines de gènes paraît moins extraordinaire, dicté par l'efficacité reproductive (ainsi notre ADN est constitué de 23 paires de chromosomes qui s'assemblent au moment de la méiose avant de se séparer en 2 brins distincts pour se reproduire. La surprise a été de constater que nous avions moins de 32 000 gènes, peut-être même seulement 25 000, mais, par exemple, le chromosome X a 931 gènes ce qui fait déjà 154 millions de paires de base!). Ce qui est sans doute trompeur car les mammifères ont une codification plus compacte que les autres vivants, saut évolutif bien plus tardif.
Là où les mammifères se différencient du reste des organismes, c'est dans le recours plus fréquent à l'épissage alternatif : en fonction de l'environnement cellulaire, ils fabriquent différentes combinaisons d'exons [parties codantes d'un gène séparées par des "introns" non codants qui sont éliminés] En moyenne, un gène donne naissance ainsi à 3 ARN différents. Mais certains gènes tels que le gène de la neurexine [protéines récepteurs des neurones], peuvent donner 2000 ARN distincts!
[...]
On découvre que le nombre de partenaires impliqués dans la régulation d'un gène pourrait être nettement plus important que ce que l'on pensait. Il est possible que cela explique une partie de la complexité des organismes supérieurs. Le concept de réseau au sein du génome, reposant sur de multiples boucles de rétroaction positives et négatives, réapparaît.
[...]
Il y a aussi toute une panoplie d'ARN non codants [ne fabriquant pas de protéines]. Ce qui est surprenant, c'est que ces derniers sont codés par des régions situés "entre" les gènes, zones que l'on pensait non transcrites jusqu'à présent. Par ailleurs, on a découvert que certains ARN non codants étaient produits par des gènes codant aussi des protéines. Notre ignorance à leur sujet est quasi complète.
[...]
Avec une redéfinition du gène, il pourrait donc y avoir des dizaines de milliers de gènes supplémentaires [actuellement autour de 25000]. La Recherche, octobre 2005
 Le
stade suivant de production de
protéines
à partir de l'ARN ne présente
pas
beaucoup de mystère non plus puisque c'est ainsi que les
protéines
sont formés dans les ribosomes
des cellules, par des ribozymes
qui sont des ARN
enzymes (les ribosomes sont des ribozymes!). Comment c'est
arrivé ? C'est une autre histoire ! Il est possible que ce soit
venu petit à petit, en attachant des acides aminés au
bout d'un ARN devenu ainsi plus réactif (sur le modèle
des ARN de transfert ci-contre, assurant la traduction
anticodons-acides aminés). En tout cas, avec les
protéines ce sont les fonctions de la cellule qui se
multiplient, ses capacités de réaction et de
communication, l'étendue de son vocabulaire, puisqu'on passe de
4
bases à 20 acides aminés. On peut parler désormais
d'un
véritable métabolisme et d'un saut de complexité
avec ce qui est sans doute l'ancêtre commun de toute cellule
(appelé LUCA, Last Universal Common Ancestor).
Actuellement on pense que le nombre minimum nécessaire pour un
métabolisme complet serait de l'ordre de 300 protéines
(gènes). [En fait l'hypothèse d'un ancêtre
commun à ce niveau me semble absurde car il faudrait que cette
cellule évolue très longtemps sans se diviser. Il est
plus probable qu'il y a eu une population de départ avec
échanges de gènes et sélection tardive d'une
structure très optimisée]
Le
stade suivant de production de
protéines
à partir de l'ARN ne présente
pas
beaucoup de mystère non plus puisque c'est ainsi que les
protéines
sont formés dans les ribosomes
des cellules, par des ribozymes
qui sont des ARN
enzymes (les ribosomes sont des ribozymes!). Comment c'est
arrivé ? C'est une autre histoire ! Il est possible que ce soit
venu petit à petit, en attachant des acides aminés au
bout d'un ARN devenu ainsi plus réactif (sur le modèle
des ARN de transfert ci-contre, assurant la traduction
anticodons-acides aminés). En tout cas, avec les
protéines ce sont les fonctions de la cellule qui se
multiplient, ses capacités de réaction et de
communication, l'étendue de son vocabulaire, puisqu'on passe de
4
bases à 20 acides aminés. On peut parler désormais
d'un
véritable métabolisme et d'un saut de complexité
avec ce qui est sans doute l'ancêtre commun de toute cellule
(appelé LUCA, Last Universal Common Ancestor).
Actuellement on pense que le nombre minimum nécessaire pour un
métabolisme complet serait de l'ordre de 300 protéines
(gènes). [En fait l'hypothèse d'un ancêtre
commun à ce niveau me semble absurde car il faudrait que cette
cellule évolue très longtemps sans se diviser. Il est
plus probable qu'il y a eu une population de départ avec
échanges de gènes et sélection tardive d'une
structure très optimisée]
 A partir de là, la formation
de l'ADN
par les protéines semble moins difficile qu'on pourrait le
croire sous prétexte
qu'il s'est imposé universellement après. Il suffit que
des
protéines ou un rétro-virus aient
transformé le ribose de l'ARN en désoxyribose, ensuite la
rigidité de l'ADN, sa stabilité et son caractère
inerte, lui ont donné un avantage reproductif
décisif, séparant la mémoire
génétique de la catalyse elle-même, ce que
renforcera le noyau dans un deuxième temps. Il est possible,
voire probable, que l'ADN soit apparu d'abord chez les virus (selon
Patrick Forterre, p39) mais ce n'est pas un
événement tellement improbable puisqu'il s'est
peut-être produit deux fois à peu près à la
même époque, de façon différenciée
chez les bactéries et les archéobactéries (ou
archées, bactéries des milieux extrêmes,
thermophiles, fonds marins, 10 fois plus petites que les
bactéries et sans doute plus originaires). L'apparition de l'ADN
en stabilisant le
génome, puis en substituant la thymine à l'uracile, ce
qui permettra d'éliminer une des sources d'erreur les plus
courantes (la dérive
de cytosine en uracile),
va diminuer brutalement la vitesse d'évolution des
protéines en améliorant considérablement la
fiabilité de la reproduction. C'est ce que Carl Woese appelle la
"cristallisation
génétique". On passe dès lors dans un tout autre
régime.
A partir de là, la formation
de l'ADN
par les protéines semble moins difficile qu'on pourrait le
croire sous prétexte
qu'il s'est imposé universellement après. Il suffit que
des
protéines ou un rétro-virus aient
transformé le ribose de l'ARN en désoxyribose, ensuite la
rigidité de l'ADN, sa stabilité et son caractère
inerte, lui ont donné un avantage reproductif
décisif, séparant la mémoire
génétique de la catalyse elle-même, ce que
renforcera le noyau dans un deuxième temps. Il est possible,
voire probable, que l'ADN soit apparu d'abord chez les virus (selon
Patrick Forterre, p39) mais ce n'est pas un
événement tellement improbable puisqu'il s'est
peut-être produit deux fois à peu près à la
même époque, de façon différenciée
chez les bactéries et les archéobactéries (ou
archées, bactéries des milieux extrêmes,
thermophiles, fonds marins, 10 fois plus petites que les
bactéries et sans doute plus originaires). L'apparition de l'ADN
en stabilisant le
génome, puis en substituant la thymine à l'uracile, ce
qui permettra d'éliminer une des sources d'erreur les plus
courantes (la dérive
de cytosine en uracile),
va diminuer brutalement la vitesse d'évolution des
protéines en améliorant considérablement la
fiabilité de la reproduction. C'est ce que Carl Woese appelle la
"cristallisation
génétique". On passe dès lors dans un tout autre
régime.
Le stade suivant, au moins vers 1,8 milliards d'années si on en croit les traces de stéroïdes qui en témoignent, c'est le passage des bactéries "procaryotes" (sans noyaux où l'ADN est libre) aux cellules "eucaryotes" (avec noyaux). Ce n'est pas une mince affaire, même si on peut penser que c'est le résultat d'une phagocytose d'une archéobactérie par une bactérie (ou le contraire), puis de leur symbiose, car on retrouve dans le noyau des gènes reproducteurs appartenant aux archéobactéries tandis que les gènes opérationnels du cytoplasme sont d'origine bactérienne. L'accroissement de complexité est inimaginable dans ces cellules au moins dix mille fois plus grosses que des bactéries et dont nous sommes faits. C'est sans doute une conséquence de la constitution d'un noyau, d'une nouvelle amélioration de la fiabilité de reproduction et de la division des tâches, plutôt qu'une condition préalable. La reproduction de la cellule passe d'ailleurs sous le contrôle de gênes régulateurs qui peuvent la bloquer et devient beaucoup moins rapide que pour les bactéries. Une reproduction fiabilisée jointe à une plus grande complexité va empêcher l'optimisation du génome eucaryote, encombré comme un vieux programme informatique de vestiges inactifs depuis longtemps ("introns", appelés aussi "ADN poubelle" mais qui pourrait avoir un rôle dans la conformation spatiale de l'ADN, voir Pour la Science, no334, août 2005). Cela va renforcer aussi le rôle des virus dans les mutations génétiques et favoriser sans doute l'apparition de la reproduction sexuée (vers 900 millions d'années?). Un des avantages décisifs des cellules à noyau sur les bactéries c'est leur mécanisme de digestion interne (lysosomes) alors que les bactéries déversent leurs enzymes à l'extérieur pour se nourrir. Une autre différence encore plus essentielle, c'est la bien plus grande capacité de communication avec l'extérieur à travers la membrane (récepteurs), condition de la formation d'organismes pluricellulaires (c'est d'ailleurs sans doute la plus grande porosité de la membrane qui a dû forcer les eucaryotes à protéger son ADN par une autre membrane, celle du noyau).
 Les eucaryotes vont intégrer ensuite (au moins vers 800 millions d'années) des mitochondries,
bactéries symbiotiques transformant l'oxygène
en
énergie (comburant du glucose et des acides gras), ce qui est
devenu à la fois possible et nécessaire lorsque les
cyanobactéries ont
commencé à produire de l'oxygène.
L'oxygène, sous-produit de la vie (à partir de 2,2
milliards d'années), a
été d'abord un terrible poison décimant la plupart
des
cellules qui étaient toutes forcément "anaérobies"
au départ puisque
nées dans un monde sans oxygène. L'oxygène nous
est devenu vital et nous protège des radiations grâce
à la couche d'ozone qu'il produit mais il est toujours agressif
et on sait bien que nous
avons besoin de
notre dose quotidienne d'anti-oxydants.
Cette fonction de protection de l'oxygène a d'abord
été assurée par les peroxysomes. Une lignée
d'eucaryotes intégrera aussi des cyanobactéries qui donneront les chloroplastes
permettant de
capter l'énergie solaire pour fixer le carbone, et donnant
naissance ainsi au royaume
végétal qui entretient toujours la production
d'oxygène pour les animaux qui le respirent, et produisent en
retour du gaz carbonique respiré par les plantes. Il faut
souligner que la
séparation des fonctions de production d'énergie, prises
en charge par les mitochondries, permet d'en centraliser la
régulation (par le NO) et, surtout,
libère les échanges électriques à travers
la membrane à des fins de signalisation cellulaire (permettant
une cohésion d'ensemble).
Les eucaryotes vont intégrer ensuite (au moins vers 800 millions d'années) des mitochondries,
bactéries symbiotiques transformant l'oxygène
en
énergie (comburant du glucose et des acides gras), ce qui est
devenu à la fois possible et nécessaire lorsque les
cyanobactéries ont
commencé à produire de l'oxygène.
L'oxygène, sous-produit de la vie (à partir de 2,2
milliards d'années), a
été d'abord un terrible poison décimant la plupart
des
cellules qui étaient toutes forcément "anaérobies"
au départ puisque
nées dans un monde sans oxygène. L'oxygène nous
est devenu vital et nous protège des radiations grâce
à la couche d'ozone qu'il produit mais il est toujours agressif
et on sait bien que nous
avons besoin de
notre dose quotidienne d'anti-oxydants.
Cette fonction de protection de l'oxygène a d'abord
été assurée par les peroxysomes. Une lignée
d'eucaryotes intégrera aussi des cyanobactéries qui donneront les chloroplastes
permettant de
capter l'énergie solaire pour fixer le carbone, et donnant
naissance ainsi au royaume
végétal qui entretient toujours la production
d'oxygène pour les animaux qui le respirent, et produisent en
retour du gaz carbonique respiré par les plantes. Il faut
souligner que la
séparation des fonctions de production d'énergie, prises
en charge par les mitochondries, permet d'en centraliser la
régulation (par le NO) et, surtout,
libère les échanges électriques à travers
la membrane à des fins de signalisation cellulaire (permettant
une cohésion d'ensemble).{kind=link}
{kind=link}

A partir de là, on peut dire que
l'évolution suit son cours, les
organismes coulent de source jusqu'aux origines de l'homme, passant d'un seuil de complexité au suivant.
Le passage aux organismes
pluricellulaires est inévitable
puisqu'il a eu lieu trois fois de façon indépendante avec
les animaux (700 millions), les végétaux (500 millions) et les champignons (600 millions?).
D'après Ameisen,
ce qui fait corps c'est un "module de
dépendance" associant toxine et protection. On peut penser
que c'est plutôt l'origine commune et la spécialisation
des organes qui
crée un code commun (hormones régulatrices), une
unité de réaction (système immunitaire), un
système vivant traversé
par une même information à laquelle on répond d'un
seul choeur. En effet, contrairement aux colonies de bactéries,
les organismes pluricellulaires se constituent à partir d'une
seule cellule eucaryote (oeuf) et partagent le même code
génétique, condition de la spécialisation des
organes qui n'en expriment qu'une partie grâce à un
rôle renforcé des gènes régulateurs. Nous avons donc un certain degré de
communauté avec toute forme de vie puisque nous descendons
finalement d'un ancêtre commun et d'une cellule unique.

Correspondance entre les gènes Hox de la mouche et de la souris
 En
tout cas, ce qui sera décisif c'est l'émergence des
organismes, d'abord indifférenciés (tribolites,
méduses, éponges) puis organisés par les gènes Hox
(sans doute à partir d'un ver marin annelé et
bilatéralisé comme le Néréis de la photo, si cela ne remonte pas au minuscule vernanimalcula)
séparant les parties du corps (tête, yeux, système
nerveux, pattes, tube digestif, pompe circulatoire) et donnant
naissance d'abord aux arthropodes (crustacés, araignées,
insectes). Nous partageons ces "gènes
architectes" (ou homéotiques) avec la mouche par
exemple (et une seule mutation d'un de ces gènes suffit pour que des pattes poussent sur sa tête!).
C'est cette nouvelle structure qui produira l'explosion du
cambrien,
après des épreuves répétées (Terre
gelée, réchauffement, retour glaciations) et la
multiplication des prédateurs (dans une dialectique
prédation/stratégies de défenses), inaugurant le
paléozoïque il y a 542 millions
d'années, bien avant la colonisation de la Terre par les
araignées et scorpions vers 410 millions d'années, suivis
par les amphibiens.
En
tout cas, ce qui sera décisif c'est l'émergence des
organismes, d'abord indifférenciés (tribolites,
méduses, éponges) puis organisés par les gènes Hox
(sans doute à partir d'un ver marin annelé et
bilatéralisé comme le Néréis de la photo, si cela ne remonte pas au minuscule vernanimalcula)
séparant les parties du corps (tête, yeux, système
nerveux, pattes, tube digestif, pompe circulatoire) et donnant
naissance d'abord aux arthropodes (crustacés, araignées,
insectes). Nous partageons ces "gènes
architectes" (ou homéotiques) avec la mouche par
exemple (et une seule mutation d'un de ces gènes suffit pour que des pattes poussent sur sa tête!).
C'est cette nouvelle structure qui produira l'explosion du
cambrien,
après des épreuves répétées (Terre
gelée, réchauffement, retour glaciations) et la
multiplication des prédateurs (dans une dialectique
prédation/stratégies de défenses), inaugurant le
paléozoïque il y a 542 millions
d'années, bien avant la colonisation de la Terre par les
araignées et scorpions vers 410 millions d'années, suivis
par les amphibiens.
 Il ne restera plus ensuite, à travers une série
d'extinctions
(440, 365, 250, 145, 65 millions d'année), dont la
pire aura été celle du permien (250 millions),
qu'à
développer les
neurones (liés au mouvement animal et à la
prédation) pour optimiser le traitement de l'information et
constituer
des individus capables d'apprendre, adaptables et sociaux, permettant
d'aboutir enfin à
l'espèce humaine et au langage qui sont bien le dessus du panier
de l'évolution, quoiqu'on dise, car cela ne pouvait guère
venir beaucoup plus tôt. Chaque pas compte et prend du temps. Les
extinctions de masse constituent un accélérateur au
moins, un nouveau départ qui fait place nette, remettant toutes
les espèces à égalité dans la
compétition de l'espace. Car le reste du temps, il y a rarement
compétition des espèces et encore moins de colonisation
d'espaces vierges. Beaucoup plus souvent, on a un équilibre des
populations prédateurs/proies relativement régulé
et ne laissant aucune place à de nouveaux entrants (le
prédateur protégeant son territoire des autres
prédateurs et des autres espèces. Les prédateurs
sont des marqueurs de la stabilité écologique car ils ont
besoin de la stabilité des proies. La stabilité
écologique est la condition du prédateur qui la
renforce).
Il ne restera plus ensuite, à travers une série
d'extinctions
(440, 365, 250, 145, 65 millions d'année), dont la
pire aura été celle du permien (250 millions),
qu'à
développer les
neurones (liés au mouvement animal et à la
prédation) pour optimiser le traitement de l'information et
constituer
des individus capables d'apprendre, adaptables et sociaux, permettant
d'aboutir enfin à
l'espèce humaine et au langage qui sont bien le dessus du panier
de l'évolution, quoiqu'on dise, car cela ne pouvait guère
venir beaucoup plus tôt. Chaque pas compte et prend du temps. Les
extinctions de masse constituent un accélérateur au
moins, un nouveau départ qui fait place nette, remettant toutes
les espèces à égalité dans la
compétition de l'espace. Car le reste du temps, il y a rarement
compétition des espèces et encore moins de colonisation
d'espaces vierges. Beaucoup plus souvent, on a un équilibre des
populations prédateurs/proies relativement régulé
et ne laissant aucune place à de nouveaux entrants (le
prédateur protégeant son territoire des autres
prédateurs et des autres espèces. Les prédateurs
sont des marqueurs de la stabilité écologique car ils ont
besoin de la stabilité des proies. La stabilité
écologique est la condition du prédateur qui la
renforce).
 C'est sans doute après l'explosion du Cambrien
où ils étaient encore anarchiques que les rapports
prédateurs/proies ont fini par devenir régulés
(par les couleurs notamment, mais surtout par la défense d'un
territoire),
stabilisant l'évolution et figeant les plans d'organisation du
vivant (macroévolution et biodisparité étaient
achevées à la fin du Cambrien). En effet, on peut penser
que les prédateurs sans limites (comme les humains peuvent l'être) ont fini par être
éliminés par manque de proies. Là aussi, avec le
temps, l'information a pris le pas sur la force pure (comme l'élevage a pris le pas sur la chasse).
C'est sans doute après l'explosion du Cambrien
où ils étaient encore anarchiques que les rapports
prédateurs/proies ont fini par devenir régulés
(par les couleurs notamment, mais surtout par la défense d'un
territoire),
stabilisant l'évolution et figeant les plans d'organisation du
vivant (macroévolution et biodisparité étaient
achevées à la fin du Cambrien). En effet, on peut penser
que les prédateurs sans limites (comme les humains peuvent l'être) ont fini par être
éliminés par manque de proies. Là aussi, avec le
temps, l'information a pris le pas sur la force pure (comme l'élevage a pris le pas sur la chasse).
Ce sont seulement les extinctions qui isolent d'abord, et font dériver génétiquement, puis mettent en concurrence l'adaptabilité des organismes, ce qui permet l'émergence de la complexification après chaque catastrophe. Ainsi après les amphibiens, le Carbonifère sera à partir de 360 millions d'années, la période des reptiles et des fougères. Ensuite, les premiers dinosaures apparaissent vers 280 millions d'années mais ne s'épanouiront qu'après le désastre du permien, par leur adaptation au manque d'oxygène, ne laissant place aux mammifères qu'après leur quasi disparition à la fin du Crétacé (et du secondaire), il y a 65 millions d'années.
 A chaque seuil de complexité franchi, le temps se ralentit et notamment avec
l'organisme animal, "le
champ des mutations efficaces va se rétrécissant de plus
en plus [...] A partir d'une certaine complexité, les mutations
efficaces sont presque exclusivement celles qui affectent des
gènes qui règlent la transcription d'autres gènes"
p25. Les mutations génétiques sont donc de plus en plus
pathogènes et néfastes (cancers). On peut juste
espérer par ce biais accélérer
ou ralentir, augmenter ou réduire, changer de place,
guère plus. Et encore, à condition d'avoir une isolation
des populations pendant un certain temps. L'évolution
naturelle est en bout de course,
passant le relais à l'apprentissage et aux progrès de la
technique. Nous en sommes là !
Ce n'est pourtant pas le dernier
mot de l'histoire de
la vie dont nous essayons bien imprudemment de prendre les commandes
(il n'y a pas que les OGM et le clonage. On crée des ADN avec de
nouvelles bases, certains espèrent créer des formes de
vie artificielles...). Ce n'est d'ailleurs que la suite d'une évolution humaine accélérée par une sélection culturelle
proche de la création artificielle de races animales.
"L'univers
c'est de la pensée qui émerge de la vie, qui
elle-même émerge de la matière dite inerte" Christian de Duve, La Recherche, p26.
A chaque seuil de complexité franchi, le temps se ralentit et notamment avec
l'organisme animal, "le
champ des mutations efficaces va se rétrécissant de plus
en plus [...] A partir d'une certaine complexité, les mutations
efficaces sont presque exclusivement celles qui affectent des
gènes qui règlent la transcription d'autres gènes"
p25. Les mutations génétiques sont donc de plus en plus
pathogènes et néfastes (cancers). On peut juste
espérer par ce biais accélérer
ou ralentir, augmenter ou réduire, changer de place,
guère plus. Et encore, à condition d'avoir une isolation
des populations pendant un certain temps. L'évolution
naturelle est en bout de course,
passant le relais à l'apprentissage et aux progrès de la
technique. Nous en sommes là !
Ce n'est pourtant pas le dernier
mot de l'histoire de
la vie dont nous essayons bien imprudemment de prendre les commandes
(il n'y a pas que les OGM et le clonage. On crée des ADN avec de
nouvelles bases, certains espèrent créer des formes de
vie artificielles...). Ce n'est d'ailleurs que la suite d'une évolution humaine accélérée par une sélection culturelle
proche de la création artificielle de races animales.
"L'univers
c'est de la pensée qui émerge de la vie, qui
elle-même émerge de la matière dite inerte" Christian de Duve, La Recherche, p26.
Correspondance entre les gènes Hox de la mouche et de la souris
En
tout cas, ce qui sera décisif c'est l'émergence des
organismes, d'abord indifférenciés (tribolites,
méduses, éponges) puis organisés par les gènes Hox
(sans doute à partir d'un ver marin annelé et
bilatéralisé comme le Néréis de la photo, si cela ne remonte pas au minuscule vernanimalcula)
séparant les parties du corps (tête, yeux, système
nerveux, pattes, tube digestif, pompe circulatoire) et donnant
naissance d'abord aux arthropodes (crustacés, araignées,
insectes). Nous partageons ces "gènes
architectes" (ou homéotiques) avec la mouche par
exemple (et une seule mutation d'un de ces gènes suffit pour que des pattes poussent sur sa tête!).
C'est cette nouvelle structure qui produira l'explosion du
cambrien,
après des épreuves répétées (Terre
gelée, réchauffement, retour glaciations) et la
multiplication des prédateurs (dans une dialectique
prédation/stratégies de défenses), inaugurant le
paléozoïque il y a 542 millions
d'années, bien avant la colonisation de la Terre par les
araignées et scorpions vers 410 millions d'années, suivis
par les amphibiens.{kind=link}
{kind=link} Il ne restera plus ensuite, à travers une série
d'extinctions
(440, 365, 250, 145, 65 millions d'année), dont la
pire aura été celle du permien (250 millions),
qu'à
développer les
neurones (liés au mouvement animal et à la
prédation) pour optimiser le traitement de l'information et
constituer
des individus capables d'apprendre, adaptables et sociaux, permettant
d'aboutir enfin à
l'espèce humaine et au langage qui sont bien le dessus du panier
de l'évolution, quoiqu'on dise, car cela ne pouvait guère
venir beaucoup plus tôt. Chaque pas compte et prend du temps. Les
extinctions de masse constituent un accélérateur au
moins, un nouveau départ qui fait place nette, remettant toutes
les espèces à égalité dans la
compétition de l'espace. Car le reste du temps, il y a rarement
compétition des espèces et encore moins de colonisation
d'espaces vierges. Beaucoup plus souvent, on a un équilibre des
populations prédateurs/proies relativement régulé
et ne laissant aucune place à de nouveaux entrants (le
prédateur protégeant son territoire des autres
prédateurs et des autres espèces. Les prédateurs
sont des marqueurs de la stabilité écologique car ils ont
besoin de la stabilité des proies. La stabilité
écologique est la condition du prédateur qui la
renforce).C'est sans doute après l'explosion du Cambrien
où ils étaient encore anarchiques que les rapports
prédateurs/proies ont fini par devenir régulés
(par les couleurs notamment, mais surtout par la défense d'un
territoire),
stabilisant l'évolution et figeant les plans d'organisation du
vivant (macroévolution et biodisparité étaient
achevées à la fin du Cambrien). En effet, on peut penser
que les prédateurs sans limites (comme les humains peuvent l'être) ont fini par être
éliminés par manque de proies. Là aussi, avec le
temps, l'information a pris le pas sur la force pure (comme l'élevage a pris le pas sur la chasse).
Il ne restera plus ensuite, à travers une série
d'extinctions
(440, 365, 250, 145, 65 millions d'année), dont la
pire aura été celle du permien (250 millions),
qu'à
développer les
neurones (liés au mouvement animal et à la
prédation) pour optimiser le traitement de l'information et
constituer
des individus capables d'apprendre, adaptables et sociaux, permettant
d'aboutir enfin à
l'espèce humaine et au langage qui sont bien le dessus du panier
de l'évolution, quoiqu'on dise, car cela ne pouvait guère
venir beaucoup plus tôt. Chaque pas compte et prend du temps. Les
extinctions de masse constituent un accélérateur au
moins, un nouveau départ qui fait place nette, remettant toutes
les espèces à égalité dans la
compétition de l'espace. Car le reste du temps, il y a rarement
compétition des espèces et encore moins de colonisation
d'espaces vierges. Beaucoup plus souvent, on a un équilibre des
populations prédateurs/proies relativement régulé
et ne laissant aucune place à de nouveaux entrants (le
prédateur protégeant son territoire des autres
prédateurs et des autres espèces. Les prédateurs
sont des marqueurs de la stabilité écologique car ils ont
besoin de la stabilité des proies. La stabilité
écologique est la condition du prédateur qui la
renforce).C'est sans doute après l'explosion du Cambrien
où ils étaient encore anarchiques que les rapports
prédateurs/proies ont fini par devenir régulés
(par les couleurs notamment, mais surtout par la défense d'un
territoire),
stabilisant l'évolution et figeant les plans d'organisation du
vivant (macroévolution et biodisparité étaient
achevées à la fin du Cambrien). En effet, on peut penser
que les prédateurs sans limites (comme les humains peuvent l'être) ont fini par être
éliminés par manque de proies. Là aussi, avec le
temps, l'information a pris le pas sur la force pure (comme l'élevage a pris le pas sur la chasse).Ce sont seulement les extinctions qui isolent d'abord, et font dériver génétiquement, puis mettent en concurrence l'adaptabilité des organismes, ce qui permet l'émergence de la complexification après chaque catastrophe. Ainsi après les amphibiens, le Carbonifère sera à partir de 360 millions d'années, la période des reptiles et des fougères. Ensuite, les premiers dinosaures apparaissent vers 280 millions d'années mais ne s'épanouiront qu'après le désastre du permien, par leur adaptation au manque d'oxygène, ne laissant place aux mammifères qu'après leur quasi disparition à la fin du Crétacé (et du secondaire), il y a 65 millions d'années.
A chaque seuil de complexité franchi, le temps se ralentit et notamment avec
l'organisme animal, "le
champ des mutations efficaces va se rétrécissant de plus
en plus [...] A partir d'une certaine complexité, les mutations
efficaces sont presque exclusivement celles qui affectent des
gènes qui règlent la transcription d'autres gènes"
p25. Les mutations génétiques sont donc de plus en plus
pathogènes et néfastes (cancers). On peut juste
espérer par ce biais accélérer
ou ralentir, augmenter ou réduire, changer de place,
guère plus. Et encore, à condition d'avoir une isolation
des populations pendant un certain temps. L'évolution
naturelle est en bout de course,
passant le relais à l'apprentissage et aux progrès de la
technique. Nous en sommes là !
Ce n'est pourtant pas le dernier
mot de l'histoire de
la vie dont nous essayons bien imprudemment de prendre les commandes
(il n'y a pas que les OGM et le clonage. On crée des ADN avec de
nouvelles bases, certains espèrent créer des formes de
vie artificielles...). Ce n'est d'ailleurs que la suite d'une évolution humaine accélérée par une sélection culturelle
proche de la création artificielle de races animales.
"L'univers
c'est de la pensée qui émerge de la vie, qui
elle-même émerge de la matière dite inerte" Christian de Duve, La Recherche, p26.